Vipère aspic
La Vipère aspic – Vipera aspis (Linnaeus, 1758)

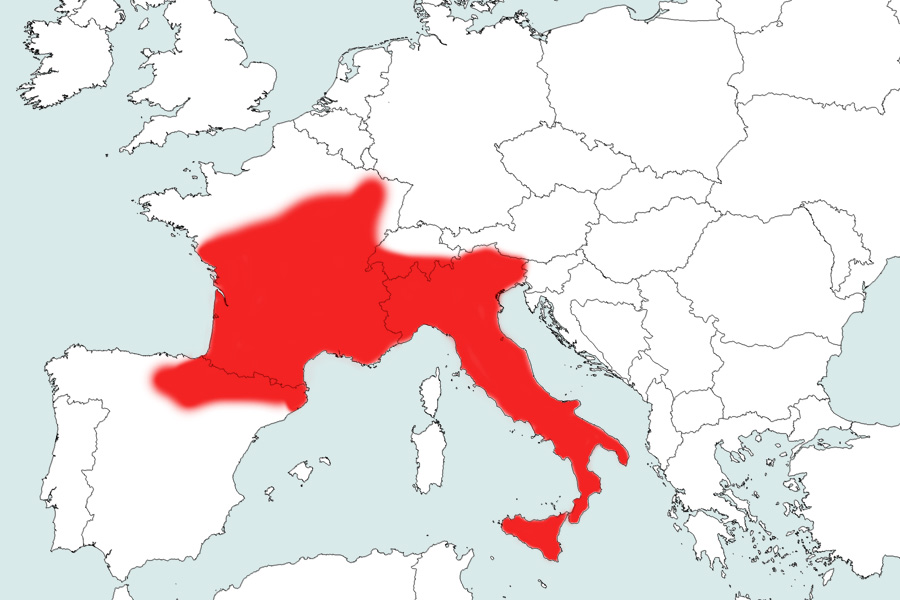
Distribution mondiale
La Vipère aspic est une espèce ouest-européenne présente en Italie (dont Sicile), en Suisse, en France (Corse exceptée), en Andorre et dans le nord-est de l’Espagne (Pyrénées et haute vallée de l’Ebre). On la rencontre ponctuellement dans l’extrême sud-ouest de l’Allemagne (Bade-Wurtemberg). L’espèce, présente dès le niveau de la mer dans de nombreuses zones, atteint des altitudes élevées sur l’ensemble des massifs montagneux de son aire de répartition : près de 3000 m dans les Alpes suisses et près de 2900 m dans les Pyrénées françaises (Saint Girons 1997, Geniez & Cheylan 2005, Gosá 2002, Hofer et al. 2001, Llorente et al. 1995, Amat Orriols & Roig Fernàndez 2003).
Distribution en France
C’est la vipère la plus largement distribuée en France et elle ne fait véritablement défaut que dans certains départements des régions du quart septentrional de l’hexagone : elle est considérée comme absente de Bretagne (mais pourrait exister dans le sud du Morbihan et l’Ile et Vilaine), elle est très localisée en Basse-Normandie (Orne et Manche) et elle est inconnue de Haute-Normandie, de Picardie et du nord de Champagne-Ardennes (Marne et Ardennes). Quelques rares populations introduites existent dans la moitié sud de l’Alsace (Haut-Rhin) (suite à l’acte illégal d’un particulier) mais l’espèce est par contre absente du Bas-Rhin.
Carte de répartition française
Variations géographiques et sous-espèces
Remarquablement polymorphe (variations entre individus au sein d’une même population) et polytypique (variations entre populations, ordonnées géographiquement), cette vipère a longtemps posé des problèmes aux systématiciens. Ainsi, Schreiber (1875) est allé jusqu’à en décrire 15 variétés ! Jusqu’à récemment, 5 sous-espèces étaient classiquement citées (Kreiner 2007, David & Vogel 2010) :
- V. a. aspis (Linnaeus, 1758) (ouest de la Suisse, France à l’exception du Sud Ouest, des Pyrénées et du sud du Massif central).
– V. a. francisciredi Laurenti, 1768 (versant méridional des Alpes centrales et Italie à l’exception de son extrémité sud).
– V. a atra Meisner, 1820 (Alpes occidentales).
– V. a. hugyi Schinz, 1834 (extrémité sud de l’Italie et Sicile).
– V. a. montecristi Mertens, 1956 (endémique de l’île italienne de Montecristo).
– V. a. zinnikeri Kramer, 1958 (sud-ouest de la France, Pyrénées et sud du Massif central).
Plusieurs travaux visant à éclaircir la phylogéographie de cette espèce ont été publiés depuis le début du 21ème siècle, dont les résultats ne sont pas tout à fait concordants (Zuffi 2002, Garrigues et al. 2005, Ursenbacher et al. 2006, Golay et al. 2008). Les deux derniers, basés sur des analyses moléculaires (mtDNA) et sur un échantillon bien réparti, invalident V. a. atra (syn. V. a. aspis) mais valident par contre V. a. francisciredi, V. a. hugyi et V. a. zinnikeri. V. a. montecristi, elle, a été placée en synonymie avec V. a. hugyi (Zuffi & Bonnet 1999, Barbanera et al. 2009).
Schématiquement, la phylogéographie moléculaire de Vipera aspis révèle l’existence de deux principales lignées évolutives : une lignée orientale correspondant aux populations d’Italie, de Sicile et du sud de la Suisse (V. aspis francisciredi et V. aspis hugyi) et une lignée occidentale correspondant à celles de la majeure partie de la Suisse, d’Allemagne, de France, d’Andorre et d’Espagne (V. aspis aspis et V. aspis zinnikeri). La relation de ces différentes lignées est la suivante au sein de l’ [espèce] :
[ (V. aspis zinnikeri – V. aspis aspis) (V. aspis francisciredi – V. aspis hugyi) ]
Soulignons que, contrairement à ce qui a été affirmé dans le passé sur la base du phénotype (et qui est encore largement relayé, à tort), la totalité du peuplement espagnol et andorran de l’espèce s’avère en fait relever, du point de vue du génotype, de la sous-espèce V. aspis zinnikeri. Nous avons donc affaire à un classique phénomène de convergence évolutive ne reflétant pas une réalité systématique (extérieurement, un dauphin ressemble plus à un requin qu’à une chauve-souris, mais il est pourtant incomparablement plus proche de la seconde).
La situation est relativement simple en France, où existent donc deux sous-lignées évolutives (V. aspis aspis et V. aspis zinnikeri) s’inscrivant toutes les deux dans la lignée occidentale. Ces deux entités sont un héritage des périodes froides du passé, qui ont durablement isolé les populations ancestrales de ces sous-espèces au sein de zones-refuge au climat plus doux (grosso modo : sud-ouest du bassin Aquitain pour les ancêtres de V. aspis zinnikeri et Midi Méditerranéen pour ceux de V. aspis aspis). Ces populations ont ensuite repris contact lors d’une extension géographique ultérieure, autorisée par une période climatique chaude.
Cet historique évolutif explique que la réalité s’avère parfois complexe sur le terrain puisque, s’agissant de sous-espèces et non pas d’espèces, V. aspis aspis et V. aspis zinnikeri se sont joyeusement métissées dans de nombreuses zones (le racisme est une tare humaine que les vipères ignorent), sur des dizaines ou centaines de kilomètres où s’observent de fascinants « dégradés » morphologiques. Dans ces endroits là, il apparaît assez vain de vouloir à tout prix attribuer à l’une ou l’autre de ces deux sous-espèces les individus observés, car ils appartiennent en réalité au deux ! Les photographies que vous pouvez observer sur notre site illustrent ce phénomène bien mieux que des mots.
En outre, comme mentionné plus haut, il existe localement des convergences évolutives qui compliquent encore la situation : des populations présentant plutôt l’aspect de V. aspis zinnikeri se révèlent appartenir à la lignée de V. aspis aspis … et inversement. Enfin, on peut trouver ça et là des populations plus ou moins différenciées du point de vue du phénotype (70% des individus avec tel ou tel motif dorsal un peu original, par ex.), qui ne présentent pourtant pas de différence significative du point de vue du génotype. C’est typiquement le cas des populations des Alpes occidentales, autrefois classées comme une sous-espèce à part (V. aspis atra) mais qui s’inscrivent totalement dans la marge de variation génotypique de la lignée V. aspis aspis. Partout, les deux sous-espèces restent très polymorphes et on n’observe jamais deux individus identiques au sein d’une population (dessin dorsal notamment).
En France, les individus les plus différenciés de V. aspis zinnikeri s’observent en Gascogne, dans deux zones distinctes : les Landes de Gascogne d’une part et la partie centrale et occidentale des Pyrénées d’autre part (du Béarn au Couserans). L’extrême sud du Massif central (Montagne Noire, Monts de Lacaune …) héberge également des animaux assez différenciés. A mesure qu’on s’éloigne de ces zones, on observe une convergence progressive vers un phénotype de plus en plus proche de celui de V. aspis aspis. La « ligne de métissage » entre les deux sous-espèces passe grossièrement par le nord de l’Aquitaine, de Midi-Pyrénées et du Languedoc-Roussillon. Au delà, le peuplement correspond à V. aspis aspis, sous-espèce qui présente localement des formes particulières : ainsi, les populations des régions méditerranéennes calcaires (« Vipère aspic de Garrigue ») possèdent invariablement une teinte de fond très pâle (souvent blanchâtre) et un motif dorsal remarquablement étroit ou quasi-indistinct.

Individus caractéristiques des sous-espèces
Vipera aspis aspis (à gauche) et Vipera aspis zinnikeri (à droite)
 Exemples de la variabilité phénotypique observée chez Vipera aspis
Exemples de la variabilité phénotypique observée chez Vipera aspis